Chlamydoselachidae is a family of primitive deep-sea sharks in the order Hexanchiformes. They are one of only two extant families in the order alongside the cow sharks in the family Hexanchidae, and the only members of the suborder Chlamydoselachoidei.
Frilled shark
The habitats of the frilled shark include the waters of the outer continental shelf and the upper-to-middle continental slope, favoring upwellings and other biologically productive areas. Usually, the shiver lives close to the ocean floor, yet its diet of cephalopods, smaller sharks, and bony fish, indicates that the frilled shark practices diel vertical migration, and swims up to feed at night at the surface of the ocean. In their Atlantic- and Pacific-ocean habitats, frilled sharks practice spatial segregation determined by the individual size, the sex, and the reproductive condition of each shark in the shiver. In Suruga Bay, on the Pacific coast of Honshu, Japan, the frilled shark is most common at the depth of 50–200 m, except in the August-to-November period, when the temperature at the 100 m water-layer exceeds 15 °C, and then the sharks swim into deeper, cooler water.
In the eastern Atlantic Ocean, the frilled shark occurs off northern Norway, northern Scotland, and western Ireland, ranging from France to Morocco, the archipelago of Madeira, and the coast of Mauritania, in northwest Africa. In the central Atlantic Ocean, the frilled shark has been caught along the region of the Mid-Atlantic Ridge, ranging from north of the Azores islands to the Rio Grande Rise, off southern Brazil, and the Vavilov Ridge, off West Africa. Frilled sharks tend to be very solitary organisms; interacting with multiple individuals of their kind is rare. However, in the late 2000s a large capture was made over an underwater seamount of the Mid-Atlantic Ridge, hauling in over 30 frilled sharks. The mass capture of a wide variety of male and female specimens emphasized these seamounts as a location for the mating of the species. In the western Atlantic, the frilled shark occurs in the waters of New England and Georgia, in the US, and in the waters of Suriname, in the northeastern coast of South America.
In the western Pacific Ocean, the frilled shark ranges from southeastern Honshu, Japan, north to Taiwan, off the coast of China, to the coast of New South Wales, Australia, and the islands of Tasmania and New Zealand. In the central and eastern Pacific Ocean, the frilled shark occurs in the regional waters of Hawaii and the coast of California, in the US, and the northern coast of Chile, in western South America. Although it has been caught at the depth of 1,570 m, the frilled shark usually does not occur deeper than 1,000 m.
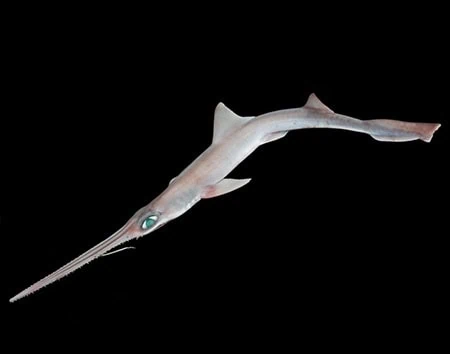
The head of the frilled shark is broad and flat, with a short, rounded snout. The nostrils are vertical slits, separated by a flap of skin that forms the incurrent opening and the excurrent opening. The moderately large eyes are horizontal ellipsoids, which have no nictitating membrane, which is a protective, third eyelid. Ligaments articulate the long jaws to the cranium, and the corners of the mouth have neither furrows nor folds. The jaws contain 300 trident-shaped teeth, each needle-tooth has a cusp and two cusplets; the rows of teeth are widely spaced, with 19–28 tooth rows in the upper jaw, and 21–29 tooth rows in the lower jaw. Frilled sharks are able to open their jaws and devour food sources that are considerably larger than their own body size, a physical trait that is also present in gulper eels and viperfish. At the throat, there are six pairs of long gill slits; the first pair of gill slits form a collar, while the extended tips of the gill filaments create a fleshy frill, hence, the frilled shark name of this fish.
The pectoral fins are short and rounded; the single, small dorsal fin has a rounded margin, and is positioned at the far end of the body, approximately opposite the anal fin. The pelvic and the anal fins are large, broad and rounded, and are positioned to the tail-end of the frilled shark’s body. The very long caudal fin is a triangular tail that has neither a lower lobe nor a ventral notch in the upper lobe, and has a margin equipped with sharp, chisel-shaped dermal denticles, which the shark can enlarge. The underside of the shark’s eel-like body features a pair of long, thick folds of skin, separated by a groove, which run the length of the belly; the function of the ventral skin-folds is unknown. In the female frilled shark, the mid-section of the body is longer, with the pelvic fins located closer to the anal fin.
A cartilaginous skeleton and a large liver (filled with low-density lipids) are the mechanical means with which the frilled shark controls and maintains its buoyancy in the deep waters of the ocean. The shark has an open, lateral-line organ system featuring mechanoreceptor hair cells in grooves exposed to the ocean environment; such a basal clade configuration enhances the frilled shark’s perception and detection of changes in the movement, the vibration, and the pressure of the surrounding water. Like all animals, the frilled shark is afflicted by parasites, such as the Monorygma tapeworm, the trematoda flatworm, the Otodistomum veliporum, and the Mooleptus rabuka nematode; and by predators, such as other sharks, as indicated by missing tail-tips lost to a hungry attacker.
The frilled shark eats a diet of cephalopods, Nudibranchs, smaller sharks, and bony fish; 60 percent of the diet is composed of squid varieties, such as the Chiroteuthis, the Histioteuthis, and the Onychoteuthis, the Sthenoteuthis and the Todarodes; and other sharks, as indicated by the stomach contents of a 1.6 m–long frilled shark which had swallowed a 590 g Japanese catshark (Apristurus japonicus). The high tendency to primarily consume the squids in their habitat can be supported by the frequent observation of beak remnants left behind during digestive processes. Because frilled sharks live on the ocean floor, they may also feed on carrion floating down from the surface.
In hunting and eating prey that are tired or exhausted or dying (after spawn), the frilled shark’s physiology suggests that it may curve its anguilline body, and brace its rear fins against the water, for leverage to effect a rapid-strike bite that captures the prey. The wide gape of the distended, long jaws allows devouring whole prey that are more than half the size of the frilled shark, itself. The jaws’ 300 recurved teeth (19–28 upper rows and 21–29 lower rows) readily snag and capture the soft body and tentacles of a cephalopod, especially with the rows of trident-shaped teeth are rotated outwards, when the jaws are open and protruded. Moreover, unlike the strong bite of sharks with an underslung jaw attached below the cranium, the frilled shark has a relatively weak bite, because of the limited leverage and force possible with long jaws that are directly articulated to the cranium, at a point behind the eyes.
The extant species of frilled shark, C. anguineus and C. africana, do not have a defined breeding season, because their oceanic habitats register no seasonal influence from the ocean’s surface; the male shark reaches sexual maturity when he is 1.0–1.2 m long, and the female shark reaches sexual maturity when she is 1.3–1.5 m long. The mature female shark has two ovaries and a uterus, which is in the right side of her body; ovulation occurs fortnightly; and pregnancy ceases vitellogenesis (yolk formation) and the production of new ova. Both ovulated eggs and early-stage shark embryos are enclosed in chondrichthyes, ellipsoid egg-cases made of a thin, golden-brown membrane.
Reproductively, the frilled shark is an ovoviviparous animal born from an encapsulated egg retained within the mother shark’s uterus. During gestation, the shark embryos develop in membranous egg-cases contained within the body of the mother shark, when the infant sharks emerge from their egg capsules in the uterus they feed on yolk until birth. The frilled-shark embryo is 3.0 cm long, has a pointed head, slightly developed jaws, nascent external gills, and possesses all fins. The growth of the jaw for elasmobranchs seem to begin early in the embryonic stage, however, it has been observed not to be the case for frilled sharks. The elongation of the jaws seemed to begin later in embryonic development. This leads to some studies suggesting that the terminal position of their mouth, due to anterior elongation of the jaw, is a derived trait instead of ancestral. When the embryo is 6–8 cm long, the mother shark expels the egg capsule, at which developmental stage the frilled shark’s external gills are developed. Throughout embryonic development, the size of the yolk sac remains constant, until the shark embryo is 40 cm long, whereupon the sac shrinks until disappearing when the embryo has grown to 50 cm in length. In the course of pregnancy, the embryo’s average rate-of-growth is 1.40 cm per month until birth, when the shark pups are 40–60 cm long, therefore, the frilled shark’s gestation period can be as long as 3.5 years; at birth, a frilled shark’s litter comprises 2–15 pups, with an average litter comprises 6.0 pups.
Southern African Frilled Shark
The southern African frilled shark is found from off the coast of southern Angola to Namibia and South Africa. Frilled sharks have also been captured off South Africa, at 1,230–1,400 m deep off Eastern Cape Province, and at 300 m deep off KwaZulu-Natal Province; it is uncertain whether these specimens are C. africana. Little is known of its habitat preferences; one known specimen was caught 425 m down in a zone of low dissolved oxygen and high nutrients, over a soft substrate.
The southern African frilled shark looks very similar to the frilled shark, with a long snake-like body and a broad, flattened head. The eyes are large and rounded. The sizable mouth is placed terminally on the blunt snout, containing around 30 tooth rows in the upper jaw and 27 tooth rows in the lower jaw. Each tooth has three slender, smooth, recurved cusps, with tiny cusplets between them, and a base that interlocks with the tooth behind it. There are six pairs of long gill slits, with the first pair meeting over the throat. The pectoral fins are broad and rounded, originating just behind the sixth gill slit. The pelvic and anal fins are large with long bases and curved margins. The single dorsal fin is set far back on the body over the anal fin, and has a short base. The caudal fin is low and somewhat triangular, without a lower lobe.
The southern African frilled shark looks very similar to the frilled shark, with a long snake-like body and a broad, flattened head. The eyes are large and rounded. The sizable mouth is placed terminally on the blunt snout, containing around 30 tooth rows in the upper jaw and 27 tooth rows in the lower jaw. Each tooth has three slender, smooth, recurved cusps, with tiny cusplets between them, and a base that interlocks with the tooth behind it. There are six pairs of long gill slits, with the first pair meeting over the throat. The pectoral fins are broad and rounded, originating just behind the sixth gill slit. The pelvic and anal fins are large with long bases and curved margins. The single dorsal fin is set far back on the body over the anal fin, and has a short base. The caudal fin is low and somewhat triangular, without a lower lobe.
Compared to the frilled shark, the southern African frilled shark has several proportional differences, including a longer head and gill slits, more widely spaced eyes and nares, broader mouth, and a greater distance between the head and the pectoral fins. The largest known female is the immature 117 cm long holotype, and the largest known males measure 99 cm long. In life the shark is dark gray, but covered with a thin membrane that gives it a uniform dark brown color.
From stomach contents, the southern African frilled shark seems to feed mainly on smaller sharks such as the African sawtail catshark (Galeus polli). Its jaws, buccal cavity, and abdomen are all highly distensible, suggesting that this shark is specialized for capturing and swallowing whole large prey, with its rows of needle-like, recurved teeth preventing escape. One 92 cm long specimen was found to have swallowed a ghost catshark (Apristurus sp.) that measured 40% of its body length.
Although adult females are unknown, the southern African frilled shark is presumed to be aplacental viviparous like the frilled shark. Males mature sexually at a length of 91.5 cm.